2.
Material und Methoden
Zur Durchführung dieser Arbeit wurde
regelmäßig eine Primärkultur von Nervenzellen aus dem Tectum
opticum des embryonalen Hühnchens angelegt. Die Zellen verschiedener
Altersstadien wurden mit Hilfe der Patch-clamp-Technik in der Whole-cell-Konfiguration
untersucht. Dabei wurden zunächst grundlegende elektrophysiologische
Eigenschaften bestimmt und dann das GABAerge Rezeptorsystem näher
charakterisiert. Die Unterscheidung von Glia- und Nervenzellen wurde anhand
elektrophysiologischer Parameter vorgenommen, eine spezielle zellbiologische
Methode wurde dafür nicht verwendet.
2.1 Anlegen von Primärkulturen
aus dem Tectum opticum
Zur Kultivierung der Zellen aus dem Tectum
opticum wurden Techniken verwendet, die eine Kultivierungsdauer bis zu
vier Wochen erlaubten (Schirm, 1993; Schlosshauer & Dütting, 1991).
Material
Geräte:
Zentrifuge
Hettich: Universal 2S, Tuttlingen
Zellzähler Casy I
Schärfe System, Reutlingen
Brutschrank
Heraeus, Hanau
Eierbrutschrank
Ehret, Emmendingen
Chemikalien:
DNAse
Gibco, Eggenstein-Leopoldshafen 1
Fötales Kälberserum
Gibco, Eggenstein-Leopoldshafen 1
Medium F12
Gibco, Eggenstein-Leopoldshafen 1
Glutamin
Gibco, Eggenstein-Leopoldshafen 1
HBSS-Puffer
Gibco, Eggenstein-Leopoldshafen 1
Hühnerserum
Gibco, Eggenstein-Leopoldshafen 1
Laminin
Sigma, Deisenhofen
Penicillin / Streptomycin
Gibco, Eggenstein-Leopoldshafen 1
(10.000 U / 100 µg / ml)
Poly-D-Lysin
Boehringer Mannheim
(1 mg / ml)
Sonstiges:
Bruteier Hisex
Weiss, Kirchberg / Iller
Deckgläser Ø 12 mm
Greiner, Nürtingen
Petrischalen Ø 60 mm
Greiner, Nürtingen
Kulturmedium:
Medium F12
+10 % Fötales Kälberserum
+2 % Hühnerserum
+2 mM Glutamin
+Penicillin (100U) / Streptomycin (100
µg / ml)
Vorbereitung
Die Kultivierung der Zellen erfolgte auf
Deckgläsern (Ø 12 mm), was für die späteren Versuche
eine einfache und flexible Handhabung erlaubte. Um eine Anheftung der Zellen
zu ermöglichen, mußten die Deckgläser folgendermaßen
vorbereitet werden:
Reinigung
Die Deckgläser wurden kurz in Aceton
getaucht, abgeflammt und dreimal durch die Flamme gezogen, um ihre Oberfläche
zu entfetten und zu reinigen. Je 5 Deckgläser wurden zusammen in eine
Petrischale (Ø 12 mm) gebracht.
Poly-D-Lysin- / Laminin-Beschichtung
Die Deckgläser wurden mit je 50/µl
Poly-D-Lysin (100/µg·ml-1)
überdeckt und eine Stunde bei 37/°C
im Brutschrank inkubiert. Anschließend wurde durch gleichzeitiges
Spülen und Absaugen mit ca. 1/ml Aqua
bidest gewaschen und eine Stunde luftgetrocknet.
Danach wurden die Deckgläser mit
je 50/µl Laminin (50/µg·ml-1)
bedeckt und über Nacht im Kühlschrank aufbewahrt.
Präparation
Die befruchteten Eier wurden von der Firma
Weiss bezogen und bei 8/°C gelagert. Die
Bebrütung erfolgte in einem Eierbrutschrank der Firma Ehret (Emmendingen)
bei 37/°C, wobei die Eier automatisch
8 mal pro Tag gewendet wurden. Zum Anlegen einer Primärkultur wurden
Eier nach dem 7. und 8. Bruttag verwendet.
Vorbereitung des Embryos
Zur Präparation wurde das Ei mit
dem stumpfen Ende nach oben in einen Eierbecher gestellt und mit 70 % Ethanol
gereinigt. Die Schale wurde mit einer Pinzette geöffnet und der Embryo
in HBSS-Puffer überführt. Daraufhin wurden Kopf- und Rumpfteil
getrennt. Das Tectum opticum war normalerweise deutlich als Wölbung
hinter dem Vorderhirn zu erkennen und konnte mit einer Pinzette leicht
abgetrennt werden. Anschließend mußten die Hirnhäute (unter
dem Binokular) entfernt werden.
Zellvereinzelung
Das gesamte Tectum wurde zur enzymatischen
Andauung in 2/ml Trypsin (1/mg·ml-1)
aufgenommen und zunächst 15/min bei Raumtemperatur,
anschließend 15/min bei 37/°C
inkubiert. Der Überstand wurde abgesaugt und das Tectum dreimal mit
kaltem HBSS-Puffer gewaschen. Das in seiner Form immer noch erhaltene Tectum
wurde nach dem Absetzen in 2/ml Kulturmedium
und 100/µl DNase (0,5/mg·ml-1)
aufgenommen und 5/min bei 37/°C
inkubiert. Anschließend wurden die Zellen durch mehrmaliges Aufsaugen
in eine Pipettenspitze mechanisch vereinzelt.
Zählung / Aussaat
Von dieser Zellsuspension wurden 1,5/ml
auf 8,5/ml Kulturmedium überschichtet
und 3/min bei 1200/U·min-1
zentrifugiert. Der Überstand wurde verworfen und das Pellet in 2/ml
Kulturmedium resuspendiert. Die Zellen wurden automatisch gezählt
(Fa. Schärfe System / Casy I) und mit einer Dichte von 100.000 bis
150.000 Zellen pro Deckglas (113/mm2)
in 50/µl Kulturmedium ausgesät.
Nach drei Stunden im Brutschrank wurden die Petrischalen mit 6/ml
Kulturmedium geflutet.
Kultivierung
Je sechs Petrischalen einer Präparation
wurden im Brutschrank bei 37/°C, 5 % CO2
und 96/% relativer Luftfeuchtigkeit bis zu
28/Tage kultiviert. Alle 2 bis 3/Tage
wurde das Kulturmedium zu 50 % ausgetauscht.
2.2 Meßtechnische Voraussetzungen
Die Patch-clamp-Technik ermöglicht
es, bei kleinen Zellen (Ø/</20/µm)
Ionenströme durch ihre Membranen zu messen. Mit einer konventionellen
Ableittechnik würden solche Zellen bei einer Penetration stark verletzt
werden, und das Signal///Rausch-Verhältnis
wäre aufgrund der zur Penetration notwendigen hochohmigen Elektrode
stark eingeschränkt.
Inzwischen gibt es eine ganze Anzahl von
Ableitkonfigurationen, die sich von der Patch-clamp-Technik herleiten (Penner,
1995). Die Voraussetzung für sämtliche Konfigurationen ist, daß
zwischen der auf die Zelle aufgesetzten Glaspipette und der Zellmembran
ein im Gigaohm-Bereich liegender Abdichtwiderstand ("GW-Seal")
zustande kommt. Die dazu notwendige Technik wurde Anfang der 80er Jahre
beschrieben (Hamill et al., 1981).
2.2.1 Patch-clamp-Messungen in der Whole-cell-Konfiguration
Spannungsklemme
In der Whole-cell-Konfiguration wird das
durch die Glaspipette eingegrenzte Membranareal zerstört und so der
elektrische Kontakt zum Zellinneren hergestellt.
Zur Messung der Membranströme muß
die Membranspannung der Zelle auf einen definierten Wert, das Haltepotential,
festgelegt werden. Der zur Einhaltung dieser Spannung notwendige Kompensationsstrom
wird von der Meßapparatur geliefert (Spannungsklemme, "voltage-clamp").
Er gibt invers die Summe der Ströme durch sämtliche geöffneten
Ionenkanäle in der Zellmembran wieder und wird als eigentlicher Meßstrom
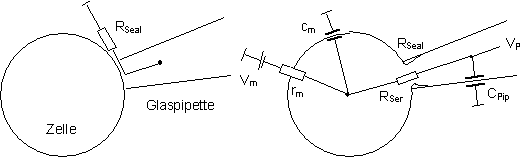
verstärkt und weiterverarbeitet (Abb. 2).
Abb. 2:
Vereinfachtes Ersatzschaltbild einer Spannungsklemme. Fließt über
die Zelle (und die Pipette) ein Strom IP, so tritt eine Differenz
zwischen dem Haltepotential VH und dem Pipettenpotential VP
auf, die an den Eingängen des Verstärkers anliegt. Dadurch wird
das Potential VV am Verstärker-ausgang geregelt, der Verstärker
fungiert als spannungsgesteuerte Spannungsquelle. VV fällt
über den Meßwiderstand R auf VP ab, um IP
nachzuliefern. Aus der Differenzspannung VA = VV
- VH läßt sich dann indirekt der Pipettenstrom IP
berechnen.
Membranströme können z.B.
durch eine Änderung des Haltepotentials ausgelöst werden, wobei
sich spannungsgesteuerte Ionenkanäle öffnen. Eine andere Möglichkeit
ist die Applikation von Substanzen, die an spezifische Rezeptoren der Zellmembran
binden und dabei einen Strom durch ligandengesteuerte Kanäle induzieren.
Die Qualität einer Spannungsklemme hängt stark von der Form der
untersuchten Zelle ab. Besitzt sie weit verzweigte Ausläufer, kann
das Haltepotential nicht an allen Membranarealen gleich gut eingehalten
werden.
Elektrische Parameter
Bei der Durchführung von Patch-clamp-Experimenten
müssen einige elektrische Parameter beachtet werden (Abb.
3). Der Abdichtwiderstand RSeal sollte mehrere GW
betragen. Der Zugriffswiderstand zur Zelle RSer muß möglichst
klein sein, um einen geringen Abfall des Signals an dieser Stelle zu gewährleisten.
RSer beträgt ca. das Doppelte des Pipettenwiderstands in
freier Badlösung.
Um Membranströme bei einer schnellen
Änderung des Membranpotentials messen zu können, müssen
die hierbei auftretenden kapazitiven Ströme, die den Verstärker
in die Sättigung treiben würden, kompensiert werden. Die wichtigsten
Quellen solcher Ströme sind die Pipettenkapazität CPip
und die Membrankapazität cm. Der Abgleich der Größen
RSer, CPip und cm kann direkt am EPC-7
Verstärker vorgenommen werden.
Abb.
3: Elektrische Parameter, die bei Patch-clamp-Messungen in der Wole-cell-Konfiguration
beachtet werden müssen: (Seal-) Abdichtwiderstand RSeal,
(Serien-) Zugriffswiderstand RSer, (Membran-) Eingangswiderstand
rm, Membranpotential Vm, Pipettenpotential VP,
Membran-kapazität cm und Pipettenkapazität CPip.
Fehlerpotentiale
An der Grenze zwischen Lösungen unterschiedlicher
Ionenzusammensetzung tritt ein "liquid-junction"-Potential auf (Barry,
1994; Ng & Barry, 1995). Da die intrazelluläre Lösung in
einer Patch-Pipette normalerweise von der Badlösung verschieden ist,
führt dies ebenfalls zu einem solchen Fehlerpotential. Bei der Korrektur
muß berücksichtigt werden, daß es nur bei der Annäherung
an eine Zelle auftritt und nach der Bildung eines Seals, und damit auch
in der Whole-cell-Konfiguration, wegfällt. Das Potential konnte für
die verwendeten Standardlösungen näherungsweise berechnet werden
und betrug ca. +4 mV. Ein mögliches Donnan-Potential im Verlauf einer
Ableitung wurde nicht korrigiert.
Material
Geräte:
Mikroskop Axiovert 35 Zeiss, Oberkochen
Verstärker EPC-7 List, Darmstadt
Oszilloskop 1602 Gould, Seligenstadt
Analogfilter npi, Tamm
Mikromanipulatoren Luigs & Neumann,
Ratingen
Piezoelement Luigs & Neumann, Ratingen
Überdruckpumpen Lorenz, Göttingen
Doppelkolbenpumpe Adams & List Assoc.,
New York
Patch-clamp-Steuerung TIDA 5.74 Battelle,
Frankfurt
DMZ Universal Puller Zeitz, Augsburg
Glaskapillaren GC150-10 Science Products,
Frankfurt
Monitor PVM 122 CE Sony, München
7-Kanal-Perfusionsnadel NMI, Reutlingen
Magnetventile Sirai, Italien
Chemikalien:
Bicucullin ((-)-Bicucullin Methchlorid)
RBI (Biotrend, Köln)
Dimethyldichlorosilan Sigma, Deisenhofen
GABA (g-Amino-Buttersäure)
Sigma, Deisenhofen
Glucose Merck, Darmstadt
Glycin Merck, Darmstadt
Kainat Sigma, Deisenhofen
Methylenchlorid Sigma, Deisenhofen
Mg-ATP Sigma, Deisenhofen
Pentobarbital Sigma, Deisenhofen
Picrotoxin RBI (Biotrend, Köln)
Strychnin Hydrochlorid RBI (Biotrend,
Köln)
Datenaufnahme und -verarbeitung:
Tida für Windows 2.63 Battelle,
Frankfurt
Sigmaplot 2.0 Jandel Corp.
Excel 5.0 Microsoft Corp.
DataCheck Eigene Programmierung mit:
Borland Pascal 7.0 Borland International,
Inc.
Zusammensetzung der verwendeten Lösungen
(Konzentrationen in mM·l-1; pH 7,4):
Standardbadpuffer
140 NaCl, 10 Glucose, 3 KCl, 1
CaCl2, 10 HEPES
Badpuffer mit reduziertem Chloridgehalt
30 NaCl, 110 Na-Propionat, 3 KCl,
1 CaCl2, 10 HEPES
Standardpipettenlösung
140 KCl, 10 HEPES, 5 EGTA, 1 CaCl2,
1 MgCl2
Pipettenlösung bei Messung des Chlorid-Umkehrpotentials
100 CsCl, 20 TEA-Cl, 10 HEPES,
5 MgCl2, 0,5 EGTA
2.2.2 Vorbereitung und Ablauf der
Messungen
Badkammer
Für die Experimente wurde jeweils
ein Deckglas mit unterschiedlich lange kultivierten Tectumneuronen in eine
Badkammer aus Plexiglas (Ø 35/mm) überführt.
Frische Badlösung wurde permanent mit einer Rate von 2/ml/·/min-1
zugeführt; mit Hilfe einer Überlaufabsaugung wurde das Badvolumen
bei ca. 1,8/ml gehalten. Daraus ergibt sich
für das Auswaschen einer Substanz eine Halbwertszeit von mindestens
45 s. Alle Messungen wurden bei Raumtemperatur durchgeführt.
Ableitelektroden
Zur Herstellung der Pipetten wurden filamentfreie
Kapillaren aus Borosilikatglas (Ø außen / innen = 1,5 / 1,05/mm)
auf einem DMZ- Universalpuller in einem dreistufigen Prozeß ausgezogen.
Die Befüllung der Spitze mit intrazellulärer Lösung erfolgte
zunächst von vorne durch Anlegen eines Unterdrucks, dann von hinten
über einen Microloader. Anschließend wurde die Pipettenspitze
in Dimethyldichlorosilan getaucht (2 % in Methylenchlorid), um die Bildung
eines Gigaseals zu erleichtern.
Über einen Silberdraht, der bei 1
- 1,5/V in einer 1/M
KCl-Lösung chloriert wurde, wurde die Elektrolytlösung in den
Pipetten mit den nachfolgenden Wandler- und Verstärkerschaltkreisen
verbunden. Als Referenzelektrode diente ein direkt in den Badpuffer eingetauchter
chlorierter Silberdraht. Bei den Messungen wird der in den Lösungen
durch Chloridionen getragene Strom in einen Elektronenstrom umgewandelt,
gemäß der Gleichung: 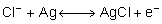 .
.
Die Osmolarität der intra- und extrazellulären
Lösungen wurde mit einem Osmometer kontrolliert (310 mosm), um ein
Anschwellen oder Schrumpfen der Zellen verhindern zu können.
Herstellung der Whole-cell-Konfiguration
Die Pipette wurde über einen Teflonhalter
direkt am Vorverstärker befestigt, der auf einem piezoelektrischen
Mikromanipulator montiert war. Um eine Verunreinigung der Pipettenspitze
zu vermeiden, wurde vor dem Eintauchen in die Badlösung ein Überdruck
von ca. 7/cm H2O angelegt. Sie
wurde durch Grobtriebe manuell in die Nähe der Zelle gebracht und
anschließend piezoelektrisch an die Zelloberfläche herangeführt
(beide Manipulatoren 3-achsig). Der Pipettenwiderstand betrug in freier
Badlösung 1 bis 3 MW.
Kurz vor dem Aufsetzen auf die Membran erhöhte sich der Pipettenwiderstand
um 20-50 %, dies wurde durch fortlaufend an der Pipette angelegte negative
Rechteckspannungen von 2/mV Amplitude sichtbar
gemacht und am Oszilloskop mitverfolgt. Hierauf wurde der Überdruck
langsam durch einen Saugdruck von 5 bis 10/cm
H2O ersetzt, was in über 90 % der Fälle zu einem Abdichtwiderstand
RSeal > 5/GW
führte. Nach Erreichen dieser als "cell-attached" bezeichneten Konfiguration
wurde zunächst ein Haltepotential von -70/mV
eingestellt und dann durch wiederholte starke Saugstöße das
unter der Pipettenspitze liegende Membranareal zerstört.
Meßdatenaufnahme
Die Meßdaten wurden durch einen
Analog / Digital-Wandler mit einer Auflösung von 16 Bit (4096 Punkte
im Meßbereich) digitalisiert und vom gleichen Computer aufgezeichnet,
der auch die Steuerung der Substanzzugabe übernahm. Die Digitalisierungsrate
betrug bei den Messungen der ligandenaktivierten Whole-cell-Ströme
400/Hz. Bei den depolarisierenden Spannungssprüngen
zur Identifizierung der Nervenzellen und bei den Messungen des Membranpotentials
betrug sie 20/kHz.
Bei sämtlichen Messungen wurde zwischen
Verstärker und dem Analog / Digital-Wandler des aufnehmenden Computers
ein Analogfilter (8-Pol Bessel, Abfall 48/dB///Oktave)
geschaltet.
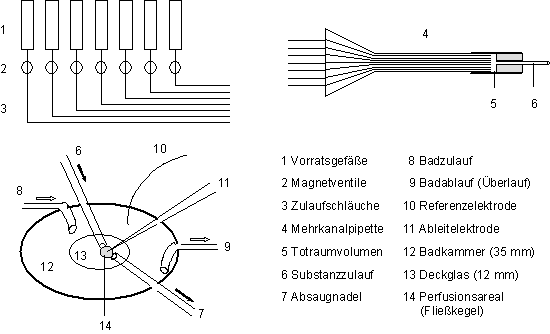
2.2.3 Perfusionssystem für Testsubstanzen
Die Testsubstanzen wurden über eine
am NMI entwickelte computergesteuerte 7-Kanal-Perfusionsnadel (Stahl, 1995)
an die jeweils zu messende Zelle herangeführt (Abb.
4). Sie wurde mit Hilfe von Mikromanipulatoren in einem Abstand von
ca. 100/µm vor der untersuchten Zelle
positioniert. Eine Absaugnadel zur schnellen Entfernung der Testsubstanzen
befand sich 200-300/µm hinter der Zelle.
Zwischen diesem Zu- und Ablauf bildete sich ein eng umgrenzter Fließkegel,
dessen Ausdehnung durch den Zusatz von Farbstoffen kontrolliert werden
konnte. Dies wurde jedoch nur zu Beginn der Untersuchungen gemacht, da
die räumliche Konfiguration des Zu- und Ablaufs bei jedem Experiment
gleich eingestellt wurde und damit auch der Fließkegel praktisch
immer die gleiche Form hatte. Durch diese Anordung konnte eine weiträumige
Kontamination der Badkammer mit Testlösungen verhindert und eine Zellcharge
über einen längeren Zeitraum verwendet werden.
Abb.
4: Schema der Meßkonfiguration und des Perfusionssystems für
Testsubstanzen.
Die Substanzaustauschzeiten dieses
Systems wurden über Änderungen des "liquid-junction"-Potentials
bei variabler Ionenzusammensetzung der Perfusionslösung getestet und
lagen bei 1-5/ms. Die "optimierte Perfusion"
(Stahl, 1995) mit Austauschzeiten unter 1/ms
wurde nicht eingesetzt, da einerseits ein zusätzlicher Kanal für
die Absaugung benötigt worden wäre und andererseits die konventionell
erreichte Anflutungsgeschwindigkeit für die geplanten Messungen ausreichend
war. Dies konnte durch einen Vergleich der Zellmembranströme gezeigt
werden, die von unterschiedlichen Substanzen ausgelöst wurden. Als
Vergleichskriterium dienten vor allem die unter Standardbedingungen durch
AMPA und Kainat induzierten Ströme.
Die Substanzzugabe erfolgte zunächst
über Schwerkraft und wurde durch Magnetventile gesteuert. Diese konnten
über ein Reizprogramm innerhalb des Meßdatenaufnahmeprogramms
zu genau definierten Zeiten betätigt werden. Später war es möglich,
auf die Vorratsgefäße der Substanzen einen Druck von bis zu
100/cm H2O zu geben, um ein konstantes
Anfluten zu gewährleisten.
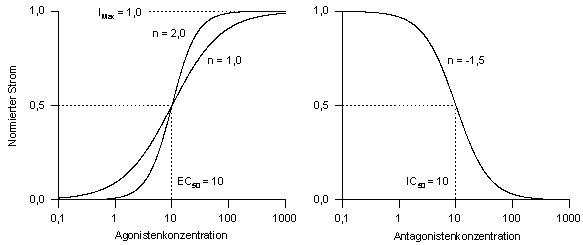
2.4 Datenauswertung
2.4.1 Dosis-Wirkungs-Kurven
Die Abhängigkeit der Amplitude des
Membranstroms von der Agonisten- (z.B. GABA, Glycin) bzw. Antagonistenkonzentration
wurde mit Hilfe von Dosis-Wirkungs-Kurven untersucht. Hierzu werden bei
unterschiedlichen Konzentrationen die Amplituden der induzierten Ströme
bestimmt und die Werte durch eine logistische Funktion der folgenden Form
angenähert:
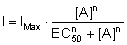 .
(1)
.
(1)
IMax bezeichnet den maximal
induzierbaren Strom, [A] die verwendete Agonisten- bzw. Antagonistenkonzentration
und n den Hillkoeffizienten als Maß der Kurvensteigung. EC50
ist bei einer Agonisten-Dosis-Wirkungs-Kurve die Konzentration, bei der
50 % des maximalen Stromes erreicht wird (Abb. 5).
Um die Messungen von Zellen mit verschiedenen absoluten Amplituden vergleichen
zu können, werden die Ströme I und IMax auf den bei
einer sättigenden Konzentration gemessenen maximalen Strom normiert
(bei GABA ³/200/µM,
bei Glycin ³/400/µM).
Zur Bestimmung der gesuchten Parameter IMax, EC50
und n stand das Programm SigmaPlot zur Verfügung. Bei der Dosis-Wirkungs-Kurve
eines Antagonisten, die auch als Hemmkurve bezeichnet und bei einer konstanten
Agonistenkonzentration gemessen wird, kann entsprechend dem Parameter EC50
in Gl. (1) ein IC50-Wert ermittelt werden. Die Steigung n der
Kurve ist in diesem Fall negativ.
Abb.
5: Veranschaulichung der verschiedenen Parameter von Dosis-Wirkungs-Kurven:
obere Asymptote IMax, Steigung n, halbmaximal effektive Konzentrationen
EC50 bzw. IC50. Die Hemmkurve bezieht sich auf einen
Strom, der durch eine konstante Agonisten-konzentration hervorgerufen wird.
Verschiedene Agonisten bzw. Antagonisten
binden an einen Rezeptor mit unterschiedlicher Affinität; ein Maß
dafür sind die Affinitätskonstanten Kd bzw. Ki.
Bei einer Agonistenkonzentration von Kd bzw. einer Antagonistenkonzentration
von Ki ist die Hälfte der Rezeptoren durch die betreffende
Substanz gebunden. Wenn dabei auch die halbmaximale Reaktion der Zelle
beobachtet werden kann, ist der Kd-Wert (Ki) mit
dem EC50-Wert (IC50) identisch, was nicht immer der
Fall sein muß.
Können jedoch Kd und EC50
gleichgesetzt werden, ergibt sich Kd sofort aus der Bestimmung
einer einfachen Agonisten-Dosis-Wirkungs-Kurve. Ki kann nicht
auf eine ähnliche Weise direkt aus dem IC50-Wert bestimmt
werden, da die Antagonistenwirkung nur in Zusammenhang mit dem Agonisten
untersucht werden kann. Der IC50-Wert ist - im Gegensatz zu
Ki - von der verwendeten Agonistenkonzentration abhängig.
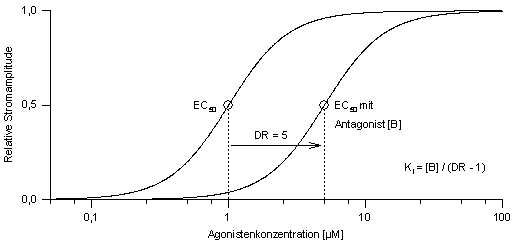
2.4.2 Hemmstoffe
Die umfassendste Methode zur Untersuchung
einer Antagonistenwirkung stellt die Schild-Analyse dar. Dabei werden mehrere
Dosis-Wirkungs-Kurven des Agonisten bei unterschiedlichen Antagonistenkonzentrationen
[B] bestimmt. Die EC50-Werte der nach rechts verschobenen Kurven
werden auf den EC50-Wert der Agonisten-Dosis-Wirkungs-Kurve
ohne Antagonisteneinfluß bezogen; dieser Verhältniswert wird
als dose ratio (DR) bezeichnet (Abb. 6).
Abb.
6: Der Wert der dose-ratio (DR) wird bestimmt, indem der EC50-Wert
der Dosis-Wirkungs-Kurve mit dem Antagonisten (normale Linie) durch den
EC50-Wert der unbeeinflußten Dosis-Wirkungs-Kurve (fette
Linie) geteilt wird. Die Affinitätskonstante Ki kann aus
der Antagonistenkonzentation [B] und dem DR-Wert berechnet werden.
Der Ki-Wert ist jene Konzentration,
bei der eine Verschiebung der Dosis-Wirkungs-Kurve um DR/=/2
nach rechts stattfindet, d.h., der EC50-Wert steigt auf das
Doppelte. Dies kommt auch in der Gaddum-Schild-Gleichung zum Ausdruck:
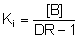 .
(2)
.
(2)
Eine Darstellung der DR-Werte in Abhängigkeit
der Antagonistenkonzentration [B] (Schild-Plot) erlaubt sowohl eine Aussage
über den Wirkungsmechanismus (kompetitiv / nicht-kompetitiv) als auch
die Bestimmung von Ki, was nur bei kompetitiven Antagonisten
sinnvoll ist.
Eine andere Untersuchungsmöglichkeit
besteht darin, die Dosis-Wirkungs-Kurve des Antagonisten (Hemmkurve, Abb.
5) bei einer festgelegten Agonistenkonzentration zu bestimmen. Dabei
werden die Amplituden bei einer gleichzeitigen Applikation des Agonisten
/ Antagonisten auf ein Kontrollexperiment ohne Antagonisten bezogen. Diese
Methode findet in solchen Fällen Anwendung, in denen aus technischen
oder zeitlichen Gründen, oder z.B. durch begrenzte Verfügbarkeit
der Substanzen, der verhältnismäßig hohe meßtechnische
Aufwand der Schild-Analyse nicht betrieben werden kann.
Dennoch kann in Fällen, in denen
ein kompetitiver Mechanismus vorausgesetzt werden darf, aus solchen Experimenten
der Ki-Wert ermittelt werden. Dazu wurde bisher häufig
eine Formel der Form
 (3)
(3)
verwendet, die ursprünglich für
Enzymbindungsstudien entwickelt wurde (Cheng & Prusoff, 1973). Hier
bezeichnet [A] die bei der Hemmkurve konstant gehaltene Agonistenkonzentration.
In jüngerer Zeit wurde gezeigt, daß bei funktionellen Experimenten
Kd durch EC50 zu ersetzen ist (Craig, 1993) und weiterhin
der Ki-Wert auch von der Steigung der Agonisten-Dosis-Wirkungs-Kurve,
also vom Hillkoeffizienten abhängt. Es ergibt sich aus diesen Überlegungen
(Leff & Dougall, 1993):
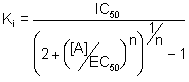 .
(4)
.
(4)
Diese modifizierte Cheng-Prusoff-Formel
wurde auch in der vorliegenden Arbeit verwendet. Neben ihr gibt es noch
weitere Methoden zur Analyse von Hemmkurven (Lazareno & Birdsall, 1993).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}